摘要:本文由365环保公司整理而成,主要讲述高钙废水颗粒污泥中古菌菌群结构变化分析,1 引言(Introduction)目前, 对颗粒污泥的研究多是关于如何快速培养、在不同进水条件(酸化与部分酸化)、不同生产工艺条件及在不同反应器中的特性等方面(郭徽等, 2018;邵享文等, 2018), 而关于含高浓度Ca2+的废纸造纸废水对生产性规模IC厌氧反应器中不同高度处颗粒污泥中古菌的影......
关键词:<strong>污水处理公司</strong>,<strong>工业水处理</strong>,<strong>污泥处理厂</strong>.
1 引言(Introduction)
目前, 对颗粒污泥的研究多是关于如何快速培养、在不同进水条件(酸化与部分酸化)、不同生产工艺条件及在不同反应器中的特性等方面(郭徽等, 2018;邵享文等, 2018), 而关于含高浓度Ca2+的废纸造纸废水对生产性规模IC厌氧反应器中不同高度处颗粒污泥中古菌的影响还鲜见报道.产甲烷菌的最大产甲烷活性是评估颗粒污泥活性最重要的指标.产甲烷菌是一个特殊的、专门的生理群, 具有特殊的细胞成分和生理功能, 是严格的专性厌氧菌, 其产甲烷途径有3种, 它们以乙酸、甲基化合物、H2/CO2为基质, 通过不同的反应途径形成甲基辅酶M, 在甲基辅酶M还原酶的催化下最终形成甲烷.产甲烷菌对pH、氧浓度的高度敏感和以H2/CO2为基质产生甲烷的自养生长, 使产甲烷菌具有繁殖世代周期长的特点.因此, 一般情况下产甲烷是有机物甲烷化过程的限速步骤.有学者在研究高浓度造纸废水厌氧处理中颗粒污泥的微生物多样性时指出, 颗粒污泥中的古菌均为甲烷菌属, 产甲烷菌包括食氢型产甲烷菌和食酸性型产甲烷菌两个生理类群(尹礎等, 2008;易敏等, 2017).此外, 邢雅娟等(2019)在研究造纸废水UMIC反应器中微生物的纵向分布特性时也发现, 系统内古菌在门分类水平上多样性较低, 主要是广古菌门和深古菌门, 在属水平上主要包括Methanosaeta、norank_p_Bathyarchaeotea、Methanobacterium、Methanosarcina等.李江等指出, 在厌氧反应器中产生的CO2在水溶液中易生成HCO3-、CO32-, 再与系统中的Ca2+生成CaCO3沉淀在颗粒污泥的内部或表面, 从而使颗粒污泥中大量的产甲烷活性受损(Yang et al., 2010;李江等, 2012).但目前还少有对CaCO3沉淀具体是影响甲烷菌的哪种类型进行分析的研究.基于此, 本文针对造纸废水中高浓度Ca2+与CO32-结合生成的CaCO3沉淀对颗粒污泥中产甲烷菌的影响进行分析, 以期能为延缓颗粒污泥的钙化提供一定的参考, 找出最适合处理废纸造纸废水的微生物种类和配比.
2 材料与方法(Materials and methods)
2.1 材料
2.1.1 厌氧颗粒污泥特性及水质分析
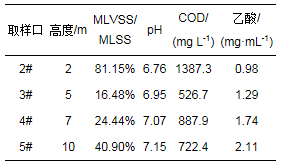
取陕西某处理废纸造纸废水的IC厌氧反应器(进水容积负荷为10~15 kg·m-3·d-1, HRT为5 h, 上升流速为5 m·h-1, 进水量为600 m3·h-1)内不同高度处的颗粒污泥(AS2), 其取样口见图 1(1#和6#取样口因冻结堵塞未能取样).种泥是河南某淀粉厂的厌氧颗粒污泥(AS1), 样品于-20 ℃的冰箱中保存待测.AS2颗粒较小, 平均粒径为0.21 mm, 呈黑灰色.厌氧进水及厌氧出水COD分别为2829、1151 mg·L-1, 去除率为59.31%.厌氧进水和出水Ca2+浓度分别为6670、3893 mg·L-1, Ca2+在厌氧反应器中的截留率为41.63%, AS2的其他特征见表 1.AS1的MLVSS/MLSS为93.46%, 颗粒较大, 平均粒径为0.35 mm, 呈青黑色空心的球形, 表面光洁.
图 1
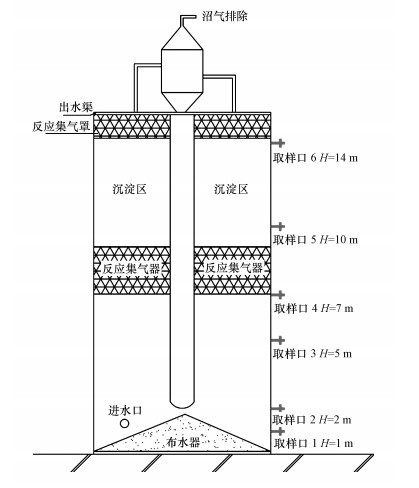
图 1 IC反应器结构示意图
表 1 AS2不同高度的污泥特征
2.1.2 实验药品及主要设备
实验药品无水乙醇、戊二醛、乙酸异戊酯、氯化钾、氯化钠、磷酸氢二钠、磷酸二氢钾、溴化钾、氯化铁、磷酸钠、无水氯化钙均为分析纯.主要仪器包括X射线衍射仪(XRD, D8 Advancee, 德国布鲁克Bruker)、红外光谱仪(IR, Vertex70, 德国布鲁克公司)、扫描电镜-能谱仪(SEM-EDX, COXEM, 捷克TESCAN)、烘箱(FCD-3000, Serials)、COD快速测定仪(5B-2F, 连华科技)、紫外-可见-近红外分光光度计(Cary 5000, 美国安捷伦).
2.2 分析方法
每个待测样品设置3个平行样, 分别进行如下的样品预处理及测定.
2.2.1 颗粒污泥基本特征及水质的测定
颗粒污泥的MLSS、MLVSS采用标准方法测定, pH使用pH计测定, COD采用连华COD快速测定仪测定, 乙酸浓度采用乙酸铁比色测定(薛超友等, 2018), 最后将各样品3个平行样的平均值作为测定结果计入表 1.
2.2.2 厌氧颗粒污泥的XRD分析
对厌氧颗粒污泥进行预处理, 每个样品的各平行样各取50 mL, 然后在105 ℃下烘干至恒重, 再将烘干后的样品置于马弗炉中在600 ℃下灼烧4 h, 灼烧完成后, 将各样品的3个平行样进行混合, 用石英研钵研细后粉体备用.利用XRD对污泥中的无机盐进行晶型分析, 波长为0.1540 nm, 管电压为40.0 kV, 管电流为40 mA, 扫描范围为10°~80°, 步长为0.02, 扫描速度正常.
2.2.3 厌氧颗粒污泥的红外光谱分析
每个样品的各平行样各取30 mL, 然后在40 ℃下烘干后将各样品的3个平行样进行混合, 研磨成粉, 最后称取0.01 g粉末样品与1 g KBr混合均匀, 倒入压片模中, 在10~80 MPa压力下加压1~2 min, 得到全透明压片.以纯KBr压片作为背景值, 在红外光谱仪中测量样品的吸光度, 测定范围为4000~500 cm-1.
2.2.4 厌氧颗粒污泥的扫描电镜-能谱分析
① 取样与清洗:每个样品的各平行样各取50 mL, 静置10 min, 去除上清液, 加入pH=7.4的磷酸盐缓冲液(PBS, 由0.24 g KH2PO4、1.44 g Na2HPO4、8 g NaCl、0.2 g KCl加蒸馏水至1 L配制而成)至原体积, 并轻微振荡混合, 冲洗3次, 以洗掉样品表面的杂质、粘液等附着物, 再静置, 自然沉淀, 去除上清液;②固定:向清洗好的颗粒污泥中加入2.5%的戊二醛溶液并淹没样品, 并置于4 ℃冰箱中固定8 h;③冲洗:固定好的颗粒污泥用PBS冲洗3次, 每次10 min;④脱水:采用梯度乙醇脱水, 将冲洗好的样品依次置于30%、50%、70%、90%、95%、100%(体积比)的乙醇中各浸泡15 min(其中, 95%和100%乙醇分别浸泡2遍);⑤置换:分别用体积比为1:1的乙酸异戊酯和乙醇的混合液及纯乙酸异戊酯各置换1次, 每次15 min;⑥干燥:将置换后的样品小心取出, 放入滤纸叠成的小盒中, 置于干燥器中干燥8 h, 干燥后将各样品的3个平行样进行混合;⑦粘样与喷金:将干燥且混合均匀的样品用镊子小心取出, 用导电胶把样品粘附在铝制托盘上, 之后在样品表面用离子溅射镀膜仪镀一层金属膜;⑧观测:利用扫描电镜扫描, 并用能谱仪分析元素组成.
2.2.5 厌氧颗粒污泥的古菌菌群生物多样性分析
将样品用冰袋冷冻运输至北京某基因科技有限公司完成PCR扩增和测序, 具体步骤如下.
① 使用PowerSoil DNA Isolation Kit(MoBio Laboratories, Carlsbad, CA)提取样品DNA, 在1%琼脂糖凝胶上检查基因组DNA的纯度和质量, 电泳条件:电压170 V, 时间30 min.然后将基因组DNA于-20 ℃下保存.
② 用引物序列为5′-ACGGGGYGCAGCAGGCGC GA-3′、5′-GGACTACVSGGGTATCTAAT-3′, 对区域16S V3~V4(344F-806R)进行PCR扩增.对每个样品, 将10位数的条形码序列添加到正向和反向引物的5′末端(由Allwegene Company, Beijing提供).使用25 μL反应液在Mastercycler Gradient(Eppendorf, Germany)上进行PCR, 其中含有12.5 μL 2×Taq PCR MasterMix、3 μL BSA(2 ng·μL-1)、2 μL引物(5 μmol·L-1)、2 μL模板DNA和5.5 μL ddH2O.PCR扩增程序为:94℃变性5 min, 35个热循环(94 ℃变性30 s, 63 ℃退火30 s, 72 ℃延伸60 s), 最后72 ℃延伸7 min.使用QIAquick凝胶提取试剂盒(QIAGEN, Germany)纯化PCR产物, 使用实时PCR定量, 并在公司进行测序.
③ 在Allwegene公司的Miseq平台上进行深度测序.运行后, 使用Illumina Analysis Pipeline Version 2.6进行图像分析、碱基调用和误差估计.
④ 通过Illumina平台进行Paired-end测序, 下机数据经过去除低质量reads(Q20, 90%标准过滤), 并剪除reads2尾部100 bp低质量序列、引物序列.
⑤ 通过COPE软件(Connecting Overlapped Pair-End, V1.2.3.3), 利用重叠关系将双末端测序得到的成对reads组装成一条序列, 利用内部编写程序去除两端barcode序列、引物序列.Paired End Reads通过reads之间的overlap(19个碱基)关系拼接成Tags, 然后去掉barcode序列.为了得到高质量的Tags, 将拼接的Tags按照长度过滤, 进行去嵌合体等处理.
3 结果分析(Results and analysis)3.1 厌氧颗粒污泥的XRD-EDS分析
如图 2a所示, AS2颗粒污泥结晶度较好, 与标准卡片谱图对比表明, 颗粒污泥中的结晶成分主要为无机盐CaCO3, 且以方解石的形式存在, 还有部分CaCO3以次稳定状态的文石存在, 含有少量的FeCO3成分.AS2中5 m处颗粒污泥中除CaCO3外还含有草酸钙和少量CaF2.如图 1b所示, AS1颗粒污泥中的无机盐主要是MgCO3、硅酸钙, 还有少量CaCO3, 峰较为宽化, 非晶相较多.
图 2
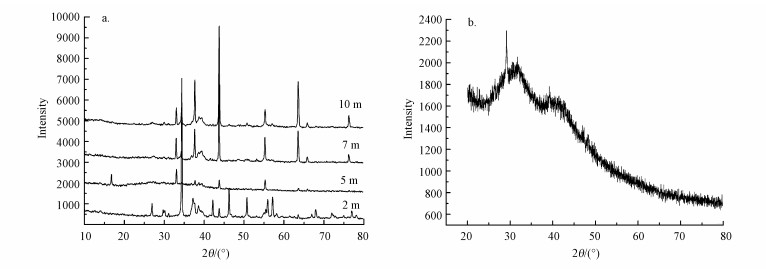
图 2厌氧颗粒污泥X-射线衍射图谱 (a.不同高度的AS2, b.AS1)
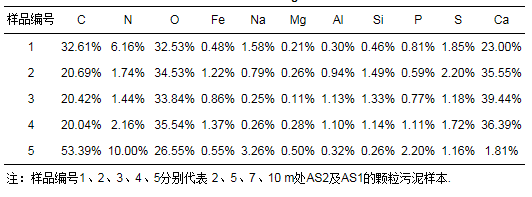
通过EDS分析检测厌氧颗粒污泥内的元素组成及比例, 结果如表 2所示.由表可知, IC反应器中AS2随着取样高度的增加, Ca含量呈现先增加后减少的变化趋势, 但5、7、10 m处颗粒污泥的Ca含量(百分比)远高于2 m处颗粒污泥, 占1/3以上, 另外含量较高的是C和O元素, Mg、Fe等其他金属元素含量较低.在AS1中Ca含量远远低于AS2中, 仅占1.81%, 含量最大的是C元素, 其次是O元素, 说明种泥中有机成分占主要地位.在AS2中, 随着取样高度的增加, 无机成分所占的比例也在增加, 这与表 1中MLVSS/MLSS的变化情况相符.结合XRD图可进一步确定AS2中Ca的主要存在形式是CaCO3, 再与AS1比较可知, 这种CaCO3的形成主要是废纸造纸废水中高浓度Ca2+与CO32-结合而成.
表 2 颗粒污泥元素
3.2 红外光谱分析
为了比较IC反应器中不同高度的颗粒污泥AS2和AS1化学结构中各官能团的分布情况, 对AS2和AS1进行了红外光谱分析(图 3).AS2中2 m和7 m处颗粒污泥在一些特定区域(3300~2850、1750~1000、750~500 cm-1)存在相同的吸收带, 但吸收峰分布有所差异.AS2中2 m处颗粒污泥在3696 cm-1处的吸收峰主要是由酸类物质的—OH伸缩振动导致(de Oliveira Silva et al., 2012;Liu et al., 2015), 这可能是因为这一区域主要发生了复杂有机物的水解酸化;各高度的AS2与AS1在3300~2850 cm-1处的吸收峰主要是由—OH和烷基上的不饱和—CH伸缩振动导致;1690~1500 cm-1处是双键的伸缩振动区;在1475~1000 cm-1处的吸收峰是X—H面内弯曲振动及X—H伸缩振动区.其中, 由于厌氧颗粒污泥成分的复杂性和蛋白质的电负性, 使AS2 4个取样口的泥样都表现为CO32-特征, 在713、700~864、844~1090、1450~1410 cm-1之间有一定的吸收峰.AS2中7 m处颗粒污泥在1407.43 cm-1处的吸收峰是矿物质中的—OH拉伸振动导致;AS2中2、5、7 m处颗粒污泥在873~707 cm-1处出现的尖锐吸收峰代表芳香族化合物, 这主要是由于废纸造纸废水成分较为复杂, 含有较多的芳香族化合物(陈永静等, 2008), 被微生物吸附富集导致.AS1与AS2的7 m处颗粒污泥呈现较为相似的红外吸收带, AS1中主要为O—H、N—H的伸缩振动和C—H的伸缩振动, 含较少的芳香族化合物, 没有明显的CO32-特征吸收峰带, 这与XRD的分析结果大体上是相符的.具体联系污水宝或参见http://www.dowater.com更多相关技术文档。
图 3

图 3不同高度的AS2(a)和AS1(b)的红外光谱
3.3 SEM分析
为了分析ICR反应器中不同高度的AS2颗粒污泥和AS1的形态结构, 采用扫描电镜(SEM)进行分析, 结果如图 4所示.AS2中2 m处的颗粒污泥内除了分布有丝状菌外, 还有少量杆菌和球菌, 同时还覆盖有大块的不规则沉淀物(图 4 a1和a2);5 m处的颗粒污泥内有大量粒状的团聚物和呈片状的沉积物, 伴随有少量的球菌及杆菌(图 4 b1和b2);7 m处的颗粒污泥内粒状团聚物和片状沉积物更加密集, 孔隙中有杆状菌存在(图 4 c1和c2);10 m处的颗粒污泥因污泥浓度较低, 样品处理后为稀少的粉末状, 但仍可见颗粒内有少量杆菌和球菌存在, 同时存在少量絮状物(图 4 d1和d2);与AS2相比, AS1则有大量规则排列的管状孔隙(图 4e1和e2), 这些孔隙就成了颗粒污泥中微生物产生的生物气和水中营养物质的传输通道, 一旦这些孔隙堵塞, 微生物的传质过程就会严重受阻, 微生物活性也因此降低.
图 4

图 4厌氧颗粒污泥的微观形态结构 (a、b、c、d与e分别为AS2的2、5、7、10 m处及AS1的颗粒污泥电镜扫描图)
3.4 古菌菌群高通量测序结果及分析
本研究利用Illumina MiSeq测序平台对AS1及AS2的2、5、7 m处的颗粒污泥样品进行测序, 结果如下.
3.4.1 Alpha多样性分析
Alpha多样性是对单个样品物种多样性的分析, 基于OTU的结果, 计算了样品的Alpha多样性(图 5).Chao1多样性估算值是根据所测的tags数和OTU数量及相对比例来预测样品中微生物的种类(OTU数量), 是基于已知结果所得的相对值.Shannon指数是一个综合OTU丰度和OTU均匀度两方面因素的多样性指数(Navarrete et al., 2015;Sengupta et al., 2015).由图 5和图 6的稀疏曲线可知, AS1的Chao1、Shannon指数、PD_whole_tree(Phylogenetic diversity)等均高于AS2, 说明种泥中物种更加丰富.稀疏曲线趋于平缓, 说明对所取样本微生物群落的检测比率接近饱和, 测序数量和测序深度都已足够反映样本的真实情况, AS2不同高度颗粒污泥中古菌的生物多样性由多到少排序为:5 m>2 m>7 m.
图 5
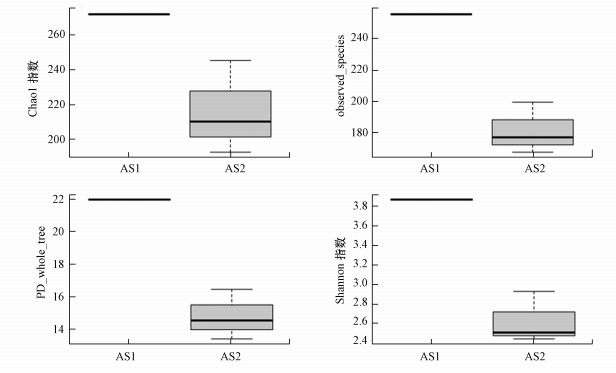
图 5 Alpha多样性分析盒型图
图 6
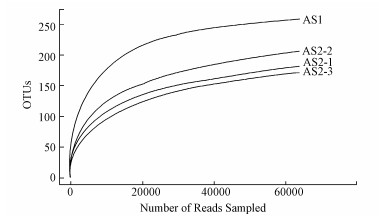
图 6 AS1和AS2样品的稀疏曲线 (AS2-1、AS2-2、AS2-3分别代表AS2的2、5、7 m处样本)
3.4.2 Beta多样性分析
在0.97的相似度下, 得到了每个样品的OTU个数.PCA分析即主成分分析, 是将多维变量降成2个变量进行分析.样本的第一主成分贡献率达79.95%, 第二主成分贡献率达18.35%, 结果显示, AS1与AS2各自聚为一类(图 7a).Flower OTU分析结果显示, 4个样本中共有95个OTU数, AS1中所特有的OTU数最多, AS2系统中5 m处样品所特有的OTU数最多, 2 m处样品次之, 7 m样品处最少(图 7b), 这与3.4.1节中的分析结果一致.
图 7
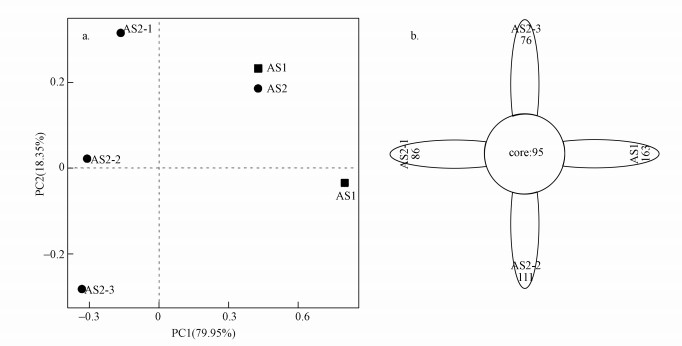
图 7 PCA分析(a)及Flower OTU分析(b)
聚类分析:基于Weighted Unifrac距离, 对样本进行聚类分析(张飞燕等, 2018;程立君等, 2019), 样本层级聚类分析结果表明(图 8), AS2-3样本与AS2-2样本聚到一起, AS2-1与AS2-2、AS2-3分开, 说明AS2中2 m处的泥样与5、7 m处的泥样有一定的差异性, 而AS1与AS2的组成差异最大.
图 8
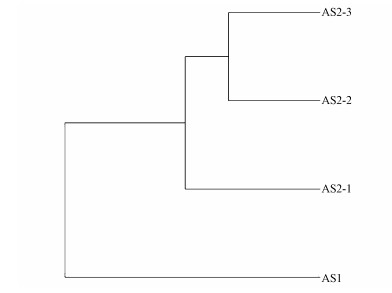
图 8样本层级聚类分析
加权UniFrac的heatmap图分析:通过对UniFrac结果的聚类, 具有相似Beta多样性的样品聚集在一起, 反映了样品间的相似性(苏瑶等, 2019), 具体结果如图 9所示.根据颜色差异可以看出样本之间的差异性, AS2中7 m处样本与5 m处样本相似性较高, 2 m处样本与5、7 m处样本具有一定的相似性, 结果与上述聚类分析的结果较为一致.
图 9
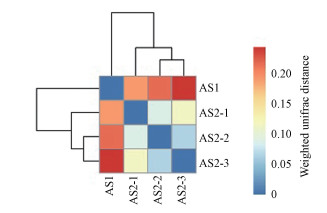
图 9加权UniFrac的heatmap
3.4.3 物种组成分析
门水平上菌群组成分析:AS1与AS2在门分类水平上的比较如图 10所示.在AS1和AS2中占比最大的门类是广古菌门, 所占比例都在90%以上;另一种主要的门类是深古菌门, 且AS1中深古菌门所占比例少于AS2中, 在AS2中随着取样高度的增加, 深古菌门所占的比例相对增加.深古菌具有多种生理生化功能, 参与甲烷代谢循环, 产乙酸, 异化还原亚硝酸盐和硫酸盐, 能降解蛋白质、多聚碳水化合物等有机质(陈玉连等, 2017).广古菌门包含了古菌中的大多数种类、产甲烷菌的全部(施嘉骏等, 2018)、极高盐度下生活的盐杆菌和一些超嗜热的好氧和厌氧菌.
图 10
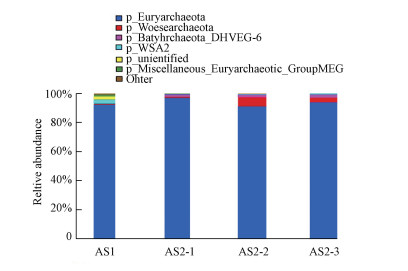
图 10门水平上的菌群组成
属水平上的菌群组成:古菌在整个微生物体系中所占的比例较小, 但对整个厌氧系统有着至关重要的作用.此处将从属分类水平上分析各样本的古菌菌群组成(图 11).在AS1中Methanosaeta(产甲烷丝菌)占42.53%, Methanolinea(产甲烷绳菌)占22.38%, 氢营养型产甲烷菌Methanoregula占13.78%, Methanobacterium(产甲烷杆菌)占6.99%, Methanospirillum(产甲烷螺菌)占2.15%, 未鉴别的和其他菌类共占12.18%.可知AS1中的优势菌分别为产甲烷丝菌、产甲烷绳菌、Methanoregula、产甲烷杆菌和产甲烷螺菌, 其相对丰度都大于1%.在AS2中2 m处的优势菌分别为产甲烷丝菌(83.22%)、产甲烷绳菌(10.81%)、产甲烷螺菌(1.38%), Methanoregula降低到0.37%;相较于AS1, 产甲烷丝菌的占比增加了近1倍, 产甲烷杆菌、产甲烷绳菌和Methanoregula等占比减少, Methanosarcina(产甲烷八叠球菌)只占0.03%.在AS2中5 m处的优势菌分别为产甲烷丝菌(85.56%)、产甲烷绳菌(2.78%), 产甲烷八叠球菌、产甲烷螺菌、Methanoregula分别占0.61%、0.45%、0.28%.在AS2中7 m处的优势菌分别为产甲烷丝菌(89.86%)、产甲烷绳菌(1.43%)、产甲烷八叠球菌(1.21%), 产甲烷杆菌、Methanoregula、产甲烷螺菌分别占0.28%、0.26%、0.15%.由图可知, 无论是在AS1中还是在AS2中最大的优势菌都是产甲烷丝菌, 但在AS2中产甲烷丝菌平均占86.21%, 相对丰度大幅度增加.
图 11
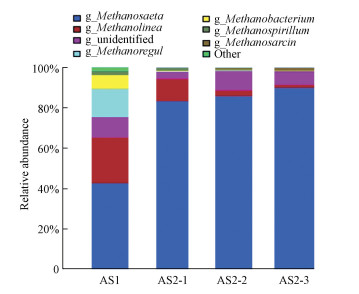
图 11属水平上的菌群组成
在AS2中随着高度的增加, 产甲烷八叠球菌和产甲烷丝菌的相对丰度呈现递增的趋势, Methanoregula、产甲烷螺菌和产甲烷绳菌则呈现递减的变化趋势.根据产甲烷菌可利用的底物类型, 可分为氢营养型、乙酸营养型和甲基营养型3种.厌氧消化过程中代谢乙酸的产甲烷菌有两类:产甲烷八叠球菌和产甲烷丝菌(杨秀山等, 1998), 产甲烷丝菌为颗粒污泥的形成提供了骨架, 从而加速了污泥的颗粒化进程.有学者认为在自然界中乙酸营养型产甲烷菌的种类较少, 只有产甲烷丝菌和产甲烷八叠球菌, 但这两种产甲烷菌在厌氧反应器中居多, 特别是前者, 因为在厌氧反应器中乙酸是主要的产甲烷基质, 一般来说, 70%左右的甲烷来自乙酸的分解.而产甲烷八叠球菌中的Methanosarcina barkeri除了以乙酸为基质外, 还能以甲醇、甲胺、H2/CO2为代谢基质(端允等, 2010;Aresta et al., 2016;王祥锟等, 2016;Batsone et al., 2016;Zabranska et al., 2017).Methanoregula、产甲烷螺菌和产甲烷杆菌是氢营养型产甲烷菌, 以H2/CO2和甲酸盐为基质, 它们能够利用H2为电子供体, CO2为电子受体, 通过电子传递还原CO2为CH4(Luo et al., 2016).由各样本的菌群组成可知, AS1中氢营养型甲烷菌和乙酸营养型甲烷菌分别占45.29%、42.6%, 在IC反应器中氢营养型甲烷菌少于17%, 乙酸营养型甲烷菌多于83.28%.可以看出, IC反应器内的CaCO3沉淀对颗粒污泥中的氢营养型甲烷菌具有较大的抑制作用.
IC反应器较大的高径比将具有不同生理生态特征的微生物分布在反应器的不同高度, 使其各自在适宜的环境条件下发挥最佳的代谢活性.有机物在厌氧消化过程主要包括水解发酵阶段、产氢产乙酸阶段、同型产乙酸阶段和产甲烷阶段.在第一阶段, 复杂有机物(如碳水化合物、蛋白质、脂肪等)转化成小分子有机物, 如脂肪酸(丙酸、丁酸等)、简单基质(甲胺、甲醇、甲酸、乙酸和H2/CO2等)及醇类物质, 两个碳以上的有机酸不能被甲烷菌直接利用, 因此, 又在产氢产乙酸菌的作用下分解成乙酸和CO2, 进而为甲烷菌提供代谢基质.班巧英等(2018)的研究表明, 氢营养型产甲烷菌比乙酸型产甲烷菌更耐酸, 氢营养型产甲烷菌主要分布在ABR反应器的1、2格室中, 乙酸营养型产甲烷菌主要分布在3、4格室.在IC反应器的下层首先进行的是水解发酵, 此时丙酸是系统中的主要有机酸, pH较低, 乙酸营养型产甲烷菌受到一定程度的抑制, 而氢营养型的产甲烷菌具有更强的耐酸能力, 能与食丙酸菌协同作用, 利用丙酸等分解的H2/CO2及甲酸甲胺等物质, 更适应这样的酸性环境.随着高度的上升, 乙酸浓度增加, 丙酸等浓度降低, 系统中pH也在增加(表 1), 生长环境和代谢基质方面都更加有利于乙酸营养型产甲烷菌的繁殖, 最终表现为随着反应器高度的增加其数量也增加.
由以上的分析结果可知, AS1中氢营养型甲烷菌占45.29%, 在IC反应器中氢营养型甲烷菌却少于17%, 氢营养型甲烷菌的相对丰度锐减, 表明CaCO3沉淀主要是对颗粒污泥中的氢营养型甲烷菌具有较大的抑制作用.氢营养甲烷菌是以H2/CO2为基质产甲烷的一类自养型产甲烷菌, 通过提高这类甲烷菌的相对比例, 从而可以减少系统中的CO2含量, 同时又能降低系统的氢分压, 最终可以促进产甲烷阶段的顺利进行, 又可以减弱系统中CO2对CO32-的贡献, 继而减少Ca2+与CO32-结合生成CaCO3沉淀在颗粒污泥上, 最终能达到延缓颗粒污泥钙化的目的.
4 结论(Conclusions)
1)在处理废纸造纸废水的生产性IC反应器内, 随着反应器高度的增加, 颗粒污泥中的无机成分也随着增加;结合XRD和EDS结果分析可知, 颗粒污泥内的无机成分主要是CaCO3, 而在种泥中的钙含量只有1.81%, 颗粒污泥的有机质含量较高.
2)颗粒污泥在高钙废水的长期作用下, 随着反应器高度的增加, 氢营养型产甲烷菌的相对丰度降低, 以Methanoregula、Methanospirillum和Methanolinea为代表, 乙酸营养型产甲烷菌的相对丰度呈现递增的趋势, 以Methanosarcina和Methanosaeta为例, Methanosaeta是各自系统中最主要的优势菌.IC反应器中不同高度处古菌的生物多样性由高到低的顺序为:5 m>2 m>7 m, 5 m处的古菌物种更加丰富.
3)AS1和AS2颗粒污泥中古菌在门分类水平上主要包括Euryarchaeota和Bathyarchaeota, 两者之和占90%以上.在属分类水平上, 主要包括Methanosaeta、Methanoregula、Methanobacterium、Methanosarcina、Methanolinea、Methanospirillum等.
4)AS1中氢营养型甲烷菌占45.29%, 在IC反应器中却少于17%, 可以看出, CaCO3沉淀使颗粒污泥的产甲烷活性降低, 主要表现为其对氢营养型甲烷菌具有较大的抑制作用.(来源:环境科学学报 作者: 张安龙)
电话
15214-307598
联系人:陈耿
邮 箱:12365@12365.co
手 机:40077-12365
地 址:上海市浦东新区张衡路200号

Copyright © 2010-2020 三六五(上海)环保有限公司 版权所有 备案号:沪ICP备19031845号-1